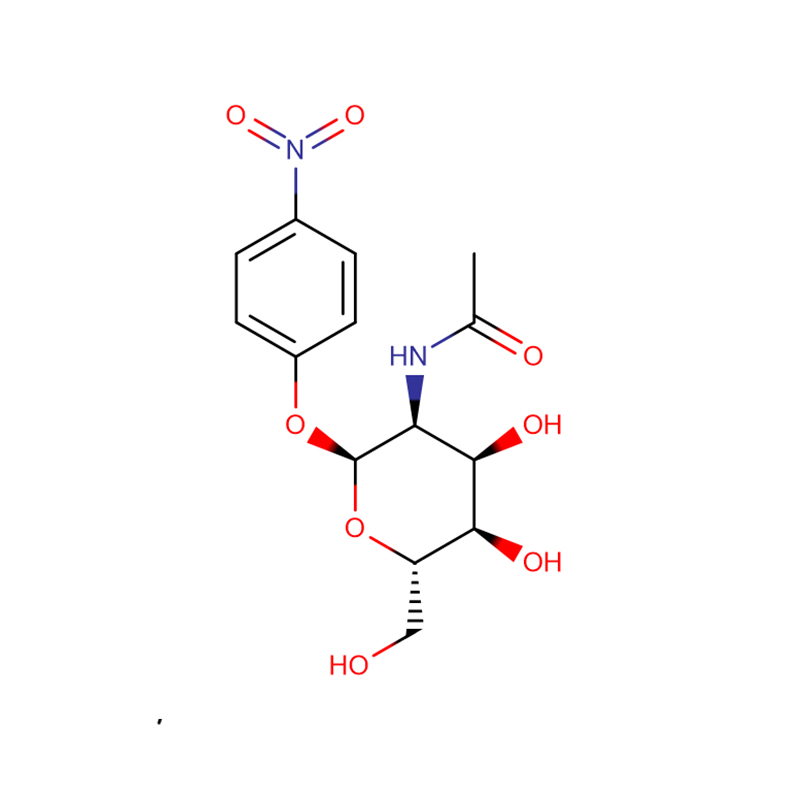
p-nitrofenylo-aL-fukopiranozyd Cas: 10231-84-2 Biały do bladożółtego krystaliczny proszek
| Numer katalogu | XD90142 |
| Nazwa produktu | p-nitrofenylo-aL-fukopiranozyd |
| CAS | 10231-84-2 |
| Formuła molekularna | C12H15NO7 |
| Waga molekularna | 285,25 |
| Szczegóły przechowywania | 2 do 8°C |
| Zharmonizowany kod taryfowy | 29400000 |
Specyfikacja produktu
| Wygląd | Krystaliczny proszek o barwie białej do bladożółtej |
| Asay | 99% |
| TLC | Pojedyncze miejsce |
| Czystość HPLC | Min. 98% |
| Rozpuszczalność (1 % w wodzie) | Klarowny bezbarwny roztwór |
| Gęstość | 1,503±0,06 g/cm3 (20ºC 760 Torr), |
| Temperatura topnienia | 196-197ºC |
| Temperatura wrzenia | 515,4°Cat760mmHg |
| Punkt zapłonu | 265,5°C |
| Współczynnik załamania światła | 1.623 |
LecA (PA-IL) jest cytotoksyczną lektyną i adhezyną wytwarzaną przez Pseudomonas aeruginosa, która wiąże hydrofobowe galaktozydy z wysoką specyficznością i powinowactwem.Wykorzystując fuzję translacyjną lecA-egfp i analizę immunoblot pozakomórkowej macierzy biofilmu, pokazujemy, że lecA ulega ekspresji w komórkach hodowanych w biofilmie.W statycznych testach biofilmu zarówno na polistyrenie, jak i stali nierdzewnej, głębokość biofilmu i pokrycie powierzchni zostały zmniejszone przez mutację lecA i wzmocnione w szczepie PAO-P47 wytwarzającym nadprodukcję LecA.Pokrycie powierzchni biofilmu przez szczep macierzysty, PAO-P47, ale nie przez mutanta lecA na stalowych kuponach, było również hamowane przez wzrost w obecności izopropylo-beta-D-tiogalaktozydu (IPTG) lub p-nitrofenylo-alfa-D-galaktozydu ( NPG).Ponadto dojrzałe biofilmy typu dzikiego utworzone przy braku tych hydrofobowych galaktozydów można zdyspergować przez dodanie IPTG.W przeciwieństwie do tego dodatek p-nitrofenylo-alfa-L-fukozy (NPF), która ma wysokie powinowactwo do lektyny P. aeruginosa LecB (PA-IIL), nie miał wpływu na tworzenie lub rozprzestrzenianie się biofilmu.Na wzrost planktonu P. aeruginosa PAO1 nie miała wpływu obecność IPTG, NPG ani NPF, a szczep nie był w stanie wykorzystać tych cukrów jako źródła węgla, co sugeruje, że obserwowany wpływ na tworzenie biofilmu wynikał z konkurencyjnego hamowania ligandu LecA wiążący.Podobne wyniki uzyskano również dla biofilmów hodowanych w warunkach dynamicznego przepływu na stalowych odcinkach, co sugeruje, że LecA przyczynia się do architektury biofilmu P. aeruginosa w różnych warunkach środowiskowych.
W liściach roślin Arabidopsis wykryto alfa-L-fukozydazę (EC 3.2.1.51) zdolną do uwalniania reszty t-fukozylowej z łańcucha bocznego oligosacharydów ksyloglukanu.Ponadto alfa-L-fukozydazę o podobnej specyficzności substratowej oczyszczono z liści kapusty (Brassica oleracea) w celu uzyskania pojedynczego prążka na SDS-PAGE.Z tego prążka białka uzyskano dwie sekwencje peptydowe i zastosowano je do identyfikacji genu Arabidopsis kodującego alfa-fukozydazę, którą proponujemy nazwać AtFXG1.Ponadto znaleziono również gen Arabidopsis wykazujący homologię ze znanymi alfa-L-fukozydazami i zaproponowaliśmy nazwanie go AtFUC1.Zarówno AtFXG1, jak i ATFUC1 ulegały heterologicznej ekspresji w komórkach Pichia pastoris, a aktywność alfa-L-fukozydazy była wydzielana do pożywki hodowlanej.Alfa-L-fukozydaza kodowana przez AtFXG1 była aktywna wobec oligosacharydów z ksyloglukanu XXFG, jak również wobec 2'-fukozylo-laktytolu, ale nie wobec p-nitrofenylo-alfa-L-fukopiranozydu.Jednakże AtFUC1 eksprymowany heterologicznie był aktywny tylko wobec 2'-fukozylo-laktytolu.Zatem to pierwsze musi być związane z metabolizmem ksyloglukanu.